 English
English 简体中文
简体中文
-

0.2ml PCR Octet Tubes
-

0.2ml PCR Octet Tubes - With Caps
-

96-Well Semi-Skirted & Non-Skirted PCR Plates
-

8-Well Enzyme Strips & Plates
-

96 Magnetic Bar Sleeve
-

8-Link Magnetic Bar Set
-
.jpg?imageView2/2/format/jp2/q/100)
96 Round Bottom Deep Well Plates
-
-1.jpg?imageView2/2/format/jp2/q/100)
96 Cone Bottom Deep Bore Plates
-

14ml 50ml Centrifuge Tubes
-

2ml 1.5ml 0.6ml Microcentrifuge Tubes


Synthesis pathway of O antigen specific polysaccharide in lipopolysaccharide
Posted by Admin | 21 Feb
Although the structure of the O antigen is diverse, its synthesis is carried out on the cytoplasmic surface. First, the sugar group is transferred to the membrane-binding lipid GCL (glycol-carrier lipid) through the nucleoside diphosphate monosaccharide precursor, and then the new O antigen is connected to the independently synthesized lipid core structure on the periplasmic surface.
1. Synthetic pathway
The O antigen synthesis pathway can be divided into three categories according to the components involved in the process of O antigen repeat unit polymerization, the location of polymerization, and the components and modes involved in the transport of polysaccharide chain across the membrane to the periplasmic space surface:
The process of O antigen repeat unit synthesis and polymerization of Salmonella typhimurium is shown in Figure 1-6.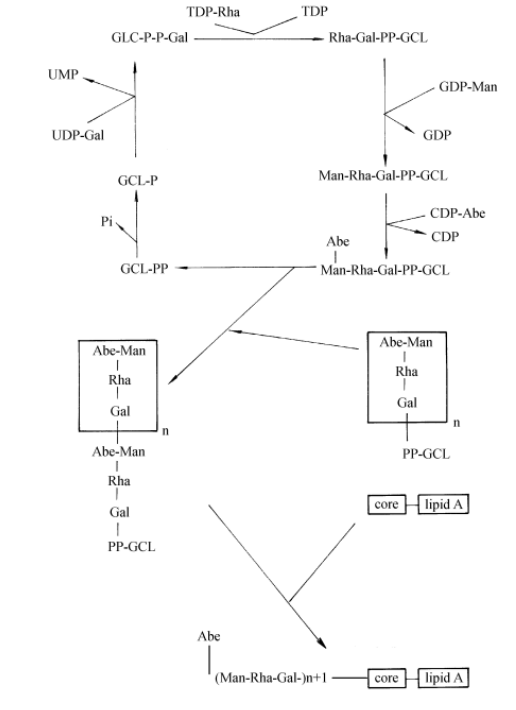
First, WbaP catalyzes the transfer of galactose 1-phosphate from UDP-Gal to GCL-P to form GCL-PP-Gal and then completes the synthesis of repeat units under the catalysis of a series of glycosyltransferases. The newly synthesized GCL-PP repeat unit is transferred from the cytoplasmic surface to the periplasmic interstitial surface, and the new GCL-PP polymer is used as the donor to transfer the polymer to the newly synthesized repeat unit. Before the next repeat unit synthesis of GCL-PP, which has hydrolyzed the polymer, GCL-P needs to be generated by pyrophosphorylase to play its role again.
For some of the O antibodies of Shigella and Escherichia coli K-12, the sugar group linked to the core polysaccharide is GlcNAc rather than Gal. The synthesis of these repeat units is initiated by UDP-GlcNA c: GCL-P-GlcNAc-1-P transferase WecA (Rfe), and the subsequent synthesis and polymerization of repeat units are the same as that of Salmonella typhimurium.
(2) ABC transporter-dependent type: this pathway is limited to the synthesis of O antigen with an extremely simple structure, which is usually a linear homomer, such as E. coli O8, O9, and Klebsiella pneumoniae O1 antigen. The synthesis is initiated by the formation of GCL-PP-GlcNAc "primer" catalyzed by WecA on the cytoplasmic surface. The glycosyltransferase continuously transfers the sugar group to the non-reducing end of the growing polymer to realize the polycondensation reaction. The polycondensation reaction does not require way's participation. The specificity of glycosyltransferase determines the structure of its repeating unit. The O antigen synthesized on the cytoplasmic surface is transferred to the periplasmic gap surface through the ABC transport device, and the connection reaction with lipid A core is completed in it.
(3) Synthase-dependent: this pathway was recently found in Salmonella O54. The structure of O54 is a poly N-acetyl mannosamine (MANNAc). Its synthesis also requires the "primer" GCL-PP-GlcNAc synthesized by WecA, and then WbbF (RfbB) catalyzes the extension of the polymer. WbbF is a progressive glycosyltransferase, or synthase, which has the dual function of transferase-output and transfers the synthesized O antigen from the cytoplasmic surface to the periplasmic surface.
Related Products
CONTACT US
Leave your name and email address, we' ll get back to you soon.
 +86-512-63121818
+86-512-63121818 +86-13036212819
+86-13036212819
+86-18018109418 [email protected]
[email protected]
[email protected] No.777, Ganquan West Road, Wujiang district, Suzhou City, Jiangsu Province, China
No.777, Ganquan West Road, Wujiang district, Suzhou City, Jiangsu Province, China
